Но если не ПД, то что?
Авторы описанных выше гипотез рассматривали механическую волну в аксоне с точки зрения термодинамики. Я же призываю взглянуть на процессы в нейроне с позиций нелинейной гидродинамики. Той самой «исконно нелинейной гидродинамики, в которой нелинейность проявляется уже в самых простых явлениях, и которая в течение почти столетия развивалась в полной изоляции от «линейной физики» [51].
А теперь, уважаемый читатель, давайте поговорим о том, о чём в нейробиологии говорить не принято.
Как в законе Ома связаны три физических величины: ток, напряжение и сопротивление (I=U/R), так же неразрывно связаны мембранный потенциал, концентрация ионов и осмотическое давление внутри клетки. Они представляют собой систему, которая всегда находится в состоянии равновесия. Изменение значения любого из этой троицы вызовет изменение остальных.
В 1884 году Анри Ле Шателье (фр. Henri Louis Le Chatelier; 1850-1936) сформулировал термодинамический принцип подвижного равновесия, позже обобщённый Карлом Брауном.
Ныне этот принцип носит имя Ле Шателье – Брауна:
если на систему, находящуюся в устойчивом равновесии, воздействовать извне, изменяя какое-либо из условий равновесия (температура, давление, концентрация, внешнее электромагнитное поле), то в системе усиливаются процессы, направленные на компенсацию внешнего воздействия.
Принцип применим к равновесию любой природы: механическому, тепловому, химическому, электрическому.
Изменения внешних условий изменяет равновесные концентрации веществ. В этом случае принято говорить о нарушении или смещении химического равновесия.
Ещё в 1908 году Нернст выявил зависимость между разностью потенциалов и ионной концентрацией. Уравнение Нернста позволяет предсказать максимальный рабочий потенциал, который может быть получен в результате электрохимического взаимодействия, при известных значениях давления и температуры.
В своё время именно по этой формуле Бернштейн теоретически рассчитал величину потенциала покоя для К+ совпавшую с экспериментально измеренным потенциалом, который составил около –70 мВ.
К сожалению Бернштейн, после публикации в 1912 году своей мембранной гипотезы прекратил дальнейшие исследования, что простительно – на тот момент ему исполнилось 70 лет. Что было тому причиной – возраст, надвигавшаяся Первая мировая война или отсутствие признания его гипотезы со стороны научного сообщества мы никогда не узна́ем.
Неизвестно и как развивались бы события продолжи он свою научную деятельность. История не знает сослагательного наклонения. Прошло время, и его последователи Ходжкин и Хаксли в 1938 году возвратились к исследованию мембранной теории, правда, несколько односторонне. Они сосредоточили своё внимание на двух элементах системы – электрическом потенциале и ионной концентрации, но игнорировали остальные – давление внутри клетки и температуру. Или не игнорировали, но приняли давление и температуру за неизменные условия среды.
Настало время восстановить справедливость, рассмотреть процесс возникновения и распространения с учётом всех составляющих.
Забегая вперёд, давайте на секундочку предположим, что нервный импульс подобен пульсовой волне, что это всплеск повышенного давления, распространяющийся по аксону. Назовём его для удобства изложения – «Нейротон».
Нейротон не является синусоидальной (гармонической) волной, а представляет собой одиночную волну. И для его описания могут быть применены соответствующие математические инструменты.
Например, его скорость в аксоне можно примерно определить с помощью формулы Моенса-Кортевега, используемой для определения скорости Пульсовой волны:
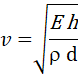
Где: Е – плотность вещества сосуда, r – модуль упругости, h – толщина клеточной мембраны, d – диаметр аксона.
r – модуль упругости – 1,05 * 103 кг/м3 (значение используется для вычисления скорости пульсовой волны).
H – толщина мембраны - 70–80 А (1А=10−10 м).
d – диаметр нервного волокна - 0.1–20 мкм.
Е – плотность вещества сосуда, для чистой воды составляет 2030 Мпа или 20300 кгс/см².
Формула Моенса-Кортевега экспериментально проверена для больших кровеносных сосудов (включая капилляры), а для таких тонких, как аксон, возможно, требуется дополнительное исследование и уточнение.
Формула применима лишь для приблизительного описания процесса распространения нейротона, поскольку объясняет распространение линейной волны, а нейротон это, возможно, нелинейная волна.
По случайному стечению обстоятельств скорость пульсовой волны примерно соответствует скорости нервного импульса в немиелинизированном нервном волокне. Достоверно известно – для людей молодого и среднего возраста скорость распространения пульсовой волны в аорте равна 5,5–8,0 м/с. С возрастом у человека уменьшается эластичность стенок артерий, и скорость пульсовой волны увеличивается. То есть применительно к нервному импульсу можно рассматривать миелиновое покрытие как «армирование» нервного волокна, которое приводит к увеличению плотности вещества сосуда и, следовательно, к увеличению скорости нервного импульса.
При прохождении нейротона по аксону происходит воздействие на мембрану (давление и растяжение), что, в свою очередь, вызывает изменение свойств мембраны, в том числе и проводимость ионов. В результате возникает хорошо всем известный электрический потенциал действия.
На приведённом рисунке изображено расширение стенок нейрона только для наглядности. Реально такого расширения может и не происходить, но это не отменяет изменения плотности внутриклеточного вещества в аксоне в момент прохождения нейротона.
И вот здесь, мы возвращаемся к нашим рассуждениям о равновесии системы из трёх составляющих – потенциал, концентрация ионов, давление (принцип Ле Шателье – Брауна). Изменяется внутренне давление – изменяется ионная концентрация, меняется потенциал на мембране. Кроме того, нейротон, проходя по аксону, формирует локальную область изменённого давления, которая, в свою очередь, создаёт условия для диффузии ионов через мембрану. После прохождения волны изменённого давления, система возвращается в исходное состояние. А мы наблюдаем перемещающийся вслед за нейротоном потенциал действия. Так просто.
А как же ионные каналы?
В модели Ходжкина – Хаксли само понятие «нервного импульса» является синонимом «потенциала действия». Я в своей модели, предполагаю, что пробегающая по аксону волна повышенного давления растягивает мембрану, что, в свою очередь, вызывает изменение свойств белковых каналов, и как следствие влияет на проводимость ионов. В результате возникает электрический потенциал действия.

Рисунок 54 Модель воротного механизма ионного канала
При этом не оспариваются известные сведения ни о составе, ни о структуре биологических мембран. Но выдвигается гипотеза о том, что в момент нарастания давления во фронте нервного импульса открываются калиевые каналы, а при снижении давления – натриевые. Ионные каналы работают по принципу «ниппель». Это гораздо проще, чем объяснение работы ионных каналов через электрические потенциалы.
Таким образом, распространяющаяся вдоль аксона волна изменения электрического потенциала является не собственно нервным импульсом, а его следствием, вторичным признаком, благодаря которому до сих пор сделаны все наблюдения за его распространением.
Причём, что примечательно, величина ПД не может быть больше определённого Бернштейном значения и полностью соответствует наблюдениям – он либо есть, либо его нет. Можно предположить, что ионные каналы срабатывают при достижении определённого уровня давления Pmin в зоне прохождения нервного импульса (или пропорционального давлению изменения концентрации ионов внутри клетки).
На рисунке проиллюстрировано, что при изменении давления в некотором диапазоне величина потенциала действия может оставаться постоянной. Правда, при этом, возможно, уменьшается длительность импульса – τ, но све́дений о таких исследованиях у меня пока нет.
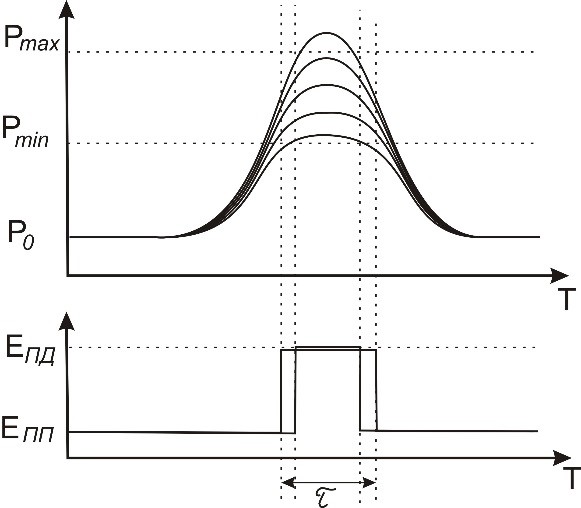
Рисунок 55. Зависимость потенциала действия от давления в аксоне.
Хотелось бы сказать, что это моё открытие, но оказалось, что всё придумано до нас, правда, применительно к системе кровообращения. Там тоже под действием изменяющегося внутреннего пульсового давления в капиллярах происходит осмотический процесс диффузии через стенку капилляра (причём в обе стороны!).
Среди гипотез, объясняющих распространение особенного (свойственного только растениям) типа мембранных потенциалов вариабельного потенциала (ВП), есть гидравлическая. Она предполагает, что ВП представляет собой местный электрический ответ, вызванный распространением гидравлической волны, которая возникает вследствие повреждающего фактора.
Данная гипотеза подтверждается обратимыми увеличениями толщины стебля, наблюдаемыми в ответ на повреждения, которые возможно, являются следствием повышения гидростатического давления в растении.
Другим аргументом в пользу гидравлического механизма служит развитие появление электрического потенциала в ответ на сжатие участка стебля (повышение давления), причём величина потенциала зависит от силы приложенного давления.
Примечательно, что скорость распространения ВП не соответствует скорости, рассчитанной с применением кабельного уравнения, что отрицает электротонический механизм его распространения. С другой стороны, не обнаружено соответствия скорости распространения ВП и скорости распространения гидравлического сигнала, индуцированного повреждением листа растения. До сих пор остаётся нерешённой проблема корреляции между относительно медленной скоростью распространения ВП и большой скоростью распространения гидравлической волны, достигающей в пределе скорости звука в водной среде (1.3 км/с). То есть, почему гидродинамическая волна распространяется так медленно?
Эта же проблема в полной мере относится и к предложенной мной гипотезе. Хотя если рассмотреть её с учётом наших знаний о распространении пульсовой волны и о солитоне, то всё, возможно, встанет на свои места.