Новейшая история нейробиологии
Вторая половина XIX века была богата открытиями в области физиологии нервных волокон, в это время были сформулированы основные законы возбуждения и распространения нервных импульсов.
Эдуард Фридрих Вильгельм Пфлюгер (Eduard Friedrich Wilhelm Pflüger; 1829–1910) в 1859 году проводя исследования действия постоянного электрического тока на нерв и мышцу обнаружил, что при замыкании цепи постоянного тока на отрицательном полюсе (катоде) возникает возбуждение, а при размыкании оно отмечается на положительном полюсе (аноде); во время прохождения тока через ткань на катоде наблюдается состояние повышенной, а на аноде – пониженной возбудимости. На основании этих исследований он сформулировал закон электротона. Учение Э. Пфлюгера об электротоне, развитое впоследствии Б. Ф. Вериго, составило основу представлении о процессах возбуждения.
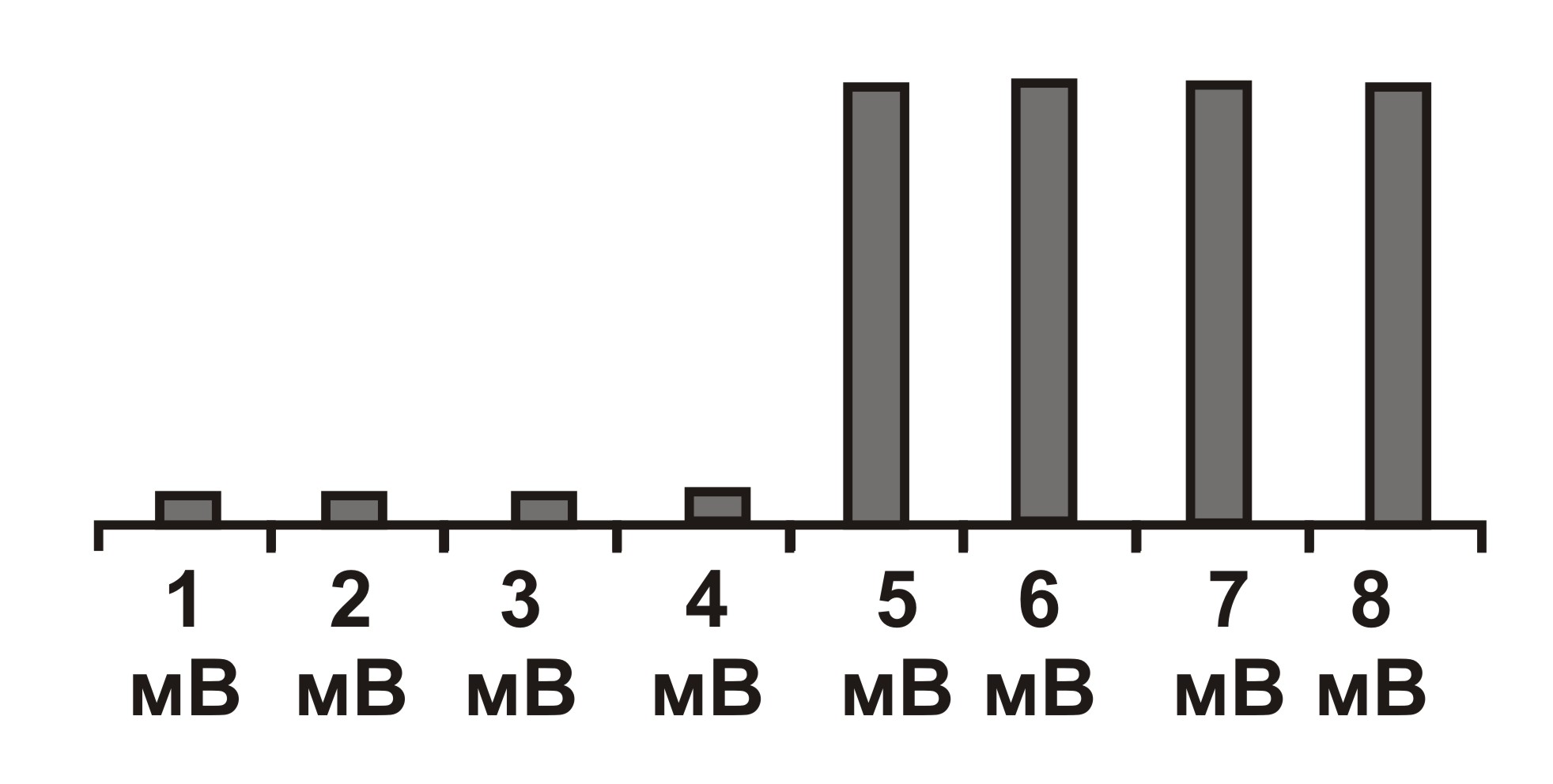
«Всё или ничего». Согласно закону Боудича (1840–1911), подпороговые раздражения не вызывают возбуждения («ничего»), при величине стимулов выше пороговых, возбуждение сразу приобретает максимальную величину («всё») и больше не возрастает при дальнейшем усилении раздражения. По этому закону функционируют и мышечные, и нервные волокна. [15]
Эта закономерность первоначально была открыта Г. Боудичем в 1876 году при исследовании сердца, а в позднее подтверждена и на всех других возбудимых тканях. Так, в 1945 году японский физиолог Генити Като́ (1890–1979) проводил эксперименты на изолированной системе. Он изолировал мышечное волокно и действовал раздражителями различной силы. В качестве раздражителя использовал электрический ток. При стимуляции импульсами подпороговой величины 1, 2, 3, 4 мВ мышечное волокно не сокращалось. А вот стимул напряжением 5 мВ приводил к сокращению изолированного волокна, при этом последующее увеличение напряжения не вызывало роста амплитуды сокращения.
В 1922-1925 годах Эдгар Дуглас Эдриан воспользовавшись капиллярным электрометром и только что изобретённым ламповым усилителем Герберта Гассера смог записать электрический потенциал отдельных нервных волокон при физическом воздействии.
Случайное наблюдение, сделанное Эдрианом в процессе эксперимента в 1928 году, ещё раз доказало наличие электричества в нервных клетках. Эдриан рассказывал: – Я разместил электроды на зрительном нерве жабы в связи с некоторыми экспериментами с сетчаткой. В комнате было почти темно, и я был озадачен, услышав повторяющиеся шумы в громкоговорителе, подключённом к усилителю . Шумы указывали на то, что имела место большая импульсная активность. Только когда я сравнил шумы с моими собственными движениями по комнате, я понял, что нахожусь в поле зрения гла́за жабы, и что он сигнализирует о том, что я делаю [16].
Примечание. Ещё в 1849 г. Дюбуа Реймон соединив роговицу и дно только что удалённого гла́за лягушки с помощью неполяризующихся электродов с гальванометром обнаружил разность потенциалов в 4—10 мВ. Так-что заслуга Эдриана не в открытии электрического потенциала в глазу земноводного, а в обнаружении корреляции между интенсивностью воздействия и частотой следования импульсов.
Эдриан подтвердил, что нервы подчиняются принципу «все или ничего». Но он также обнаружил, что применительно к нервам закон «все ли ничего» имеет продолжение: амплитуда нервных импульсов действительно сохраняется одинаковой, но при этом – с ростом силы раздражения может формироваться серия нервных импульсов, и чем сильнее раздражитель, тем больше частота их следования. Вероятно, так обеспечивается градация интенсивности ощущений. «В связи с этим импульсация несёт гораздо большую информацию, чем просто сигнал о том, что возбуждение произошло», – писал Эдриан [16].
Кроме того, он обнаружил, что более сильный стимул активирует большее количество чувствительных волокон.
Тогда же сложилось и устойчивое представление о том, что сигналы возбуждений, приходящие на разные дендриты, суммируются в соме нервной клетки и в результате формируется исходящий сигнал в аксоне.

Однако, последние исследования нейробиологов из Израиля, опубликованные в 2018 году в научном издании Scientific Reports опровергают эту модель. Получены свидетельства того, что направление результирующего сигнала существенно может повлиять на реакцию нейрона. К примеру, слабый сигнал «слева» и примерно такой же «справа» нейрон не суммирует и не отзовётся выходным импульсом, но если сигнал с бо́льшей мощностью поступит с одной из сторон, то запустить реакцию нейрона может даже он один [17].
.
.