Овершут
Дальнейшие исследования показали, что вариант с «дыркой» в мембране предложенный Ю.Бернштейном не состоятелен: возникновение потенциала действия невозможно объяснить простым «закорачиванием» мембраны. Более того, эксперименты с измерением сопротивления мембраны показали, что в реальности ПД оказался заметно больше ПП.
Надо было как-то объяснить возникновение этого «дополнительного» потенциала. На самом деле для решения этой задачи все данные были уже известны, основные идеи высказаны – оставалось только их сопоставить и сделать решающий вывод.
Суть новой гипотезы заключалась в следующем. Кроме основного потенциала покоя, создаваемого ионами калия внутри клетки, существует ещё один «встречный» потенциал, создаваемый ионами натрия, находящимися во внеклеточной среде. Эти потенциалы суммируются. Пока нейрон пребывает в состоянии покоя, потенциалзависимые каналы мембраны закрыты и поддерживается равновесный суммарный ПП. Когда в результате возбуждения мембранный потенциал увеличивается до порогового уровня, например, с -70 до -55 милливольт, первыми открываются потенциалзависимые натриевые каналы, и ионы натрия устремляются внутрь клетки, вызывая краткое, но резкое увеличение количества положительных зарядов и поднимая мембранный потенциал до +40 милливольт. В ответ на это изменение мембранного потенциала натриевые каналы, открывшись на некоторое время, закрываются, а потенциалзависимые калиевые каналы ненадолго открываются, увеличивая отток положительно заряженных ионов калия из клетки и быстро возвращая мембранный потенциал к состоянию покоя –70 милливольт.
Совсем нелегко было прийти к таким представлениям. И вот почему: диаметр иона натрия в растворе раза в полтора больше диаметра ионов калия и хлора. И совершенно необъяснимо, как бо́льший по размеру ион проходит там, где не может пройти меньший .
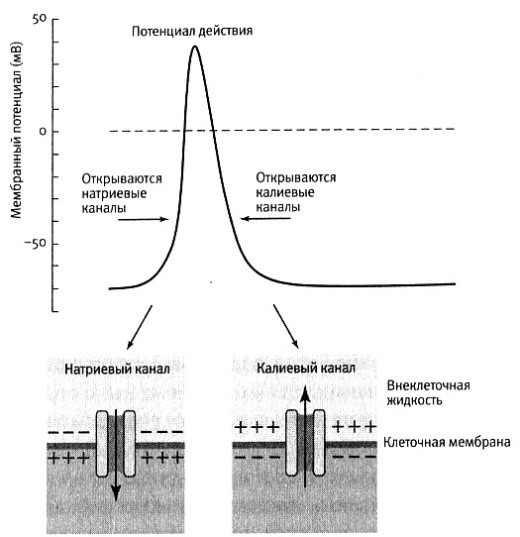
Нужно было найти новое объяснение механизма перехода ионов через мембраны. Только осмосом движение ионов через мембрану не объяснить. И тогда была высказана идея, что ионы могут пересекать мембрану с помощью пока неизвестных белковых молекул. Причём эти «молекулы-такси» хорошо различают своих пассажиров, и никогда не путают ионы натрия с ионами калия.
Так элегантно были объяснены значения потенциалов покоя и действия. А следующим шагом стало объяснение распространения нервного импульса путём последовательного открытия натриевого и калиевого каналов мембраны.
Алан Ходжкин прекрасно разбирался в электричестве и дал такую иллюстрацию распространения сигнала в нерве. «Если специалист по электричеству посмотрит на нервную систему, то сразу увидит, что передача сигнала по нервным волокнам является огромной проблемой. Диаметр аксона в нерве варьирует от 0,1 до 20 микрон. Внутреннее содержимое содержит ионы и является неплохим проводником электричества. Однако, волокно невелико и его продольное сопротивление очень большое. Простой расчёт показывает, что в волокне диаметром 1 микрон и сопротивлением 100 Ом/см удельное сопротивление составит около 1010 Ом/см. Это означает, что электрическое сопротивление маленького нервного волокна длиной в 1 метр равно сопротивлению 1010 миль 0,2 мм медной проволоки, то есть проволоки длиной в десять раз больше, чем от Земли до планеты Сатурн» [43].
Требовалось найти другое обоснование электрической природы нервного импульса. И надо сказать, он его нашёл. И, решение это, не менее элегантно, чем объяснение овершута.
Ходжкин и Хаксли совместили теорию местных токов, предложенную Лудимаром Германом, кабельную теорию Томсона и собственную мембранную теорию. Ив вот что у них получилось:
После того как потенциал действия возникает на одном участке аксона, создаваемые при этом ионные токи возбуждают соседние участки, вызывая изменение потенциала действия и на них. Происходящая в результате цепная реакция обеспечивает распространение потенциала действия по всей длине нервного волокна от места, где он был вызван первоначально, до терминалий, передающих сигнал другому нейрону (или мышечной клетке).
Возможно, такая схема и работала бы в идеальных условиях, где нет потерь, так почти бесконечно распространялась бы волна от брошенного в океан камня. В реальных же условиях существуют утечки и другие явления, приводящие к тому, что на каждом следующем участке аксона реполяризация будет уменьшаться и бегущая волна постепенно затухнет. И это не единственная проблема.
Возник ряд сложных и до поры до времени неразрешимых вопросов:
- Почему амплитуда нервного импульса не уменьшается в процессе распространения (НИ не затухает)?
- Как восстанавливается исходный потенциал на мембране после прохождения НИ?
- Как проходят ионы натрия и калия через мембрану?
В результате Ходжкин и Хаксли, предложили максимально полное на тот момент биофизическое описание потенциала действия, хотя методы исследования молекулярных механизмов нервного импульса стали доступными только в 80-х годах XX века.
Модели подобные Модели Ходжкина – Хаксли были созданы впоследствии и для других электрохимически возбуждаемых клеток – например, для сердечных миоцитов. И вот что примечательно, во многих русскоязычных источниках, все модели такого рода описывают как автоволновые процессы в активных средах. Более того пример автоволновых процессов в биологии напрямую отсылает читателей к модели Ходжкина – Хаксли.
Для того чтобы понять современное представление о распространении нервного импульса нам придётся сделать отступление и узнать кое-что про автоволновые процессы.